
Газета «Новости медицины и фармации» 16 (426) 2012
Вернуться к номеру
Почему мы не победим ВИЧ/СПИД-пандемию
Авторы: М.В. Супотницкий - кандидат биологических наук
Версия для печати

Окончание. Начало в № 15 (425)
В соответствии с представлениями о роли иммунной системы в защите макроорганизма от патогенных микроорганизмов, сложившимися в начале ХХ столетия и до сегодняшнего дня переписываемыми из учебника в учебник, комплемент должен контролировать ВИЧ-инфекцию, как и любую другую. Тем более что процесс развивается медленно, не сопровождается симптомами шока (как, например, это происходит при натуральной оспе или чуме). Но и комплемент при ВИЧ-инфекции ведет себя «не так». Плазма крови человека усиливает инфекционность ВИЧ в отношении мононуклеарных клеток и моноцит-производных макрофагов почти в 30 раз (S. Wu et al., 1995). В более детальных исследованиях установлено непосредственное связывание СI-домена gp120 ВИЧ с фактором Н комплемента (негативный регулятор активности комплемента, синтезируется макрофагом) и увеличение формирования синтиция CD4-зависимым образом (C. Pinter et al., 1995).
Приведенные данные показывают, что иммунная система человека не воспринимает ВИЧ как нечто для нее чужое и не вступает в борьбу с ним, что действительно противоречит общепринятым взглядам на функционирование его иммунной системы, однако это противоречие исчезает, если учитывать роль самих ретровирусов в ее эволюции. Но давайте посмотрим еще и на то, как функционируют так называемые антиретровирусные системы человека.
Таких систем как минимум две: AID/APOBEC и TRIM5-альфа. Они самостоятельны и не зависят ни друг от друга, ни от иммунной системы человека. Ген APOBEC/3G (apolipoprotein B mRNA-editing enzyme, catalytic polypeptide-like 3G) кодирует белок, который упаковывается в ретровирусные частицы, где дезаминирует цитозин на урацил в минус цепи вирусной ДНК в процессе обратной транскрипции. В результате чего в плюс цепи кДНК гуанозин заменяется аденином, и репликация ВИЧ останавливается (S. Wahl et al., 2006; B.G. Cullen, 2006).
TRIM5-альфа (tripartite interaction motif5-альфа) является тримерным белком и взаимодействует с гексамерным капсидом вируса через пролиновые пептидные группы, находящиеся в cis-конформации. Формирование комплекса «вирус — TRIM5-альфа» блокирует этап «раздевания» вируса и перенос его нуклеиновой кислоты в ядро клетки. Инфекция блокируется еще до обратной транскрипции вирусной РНК.
На первый взгляд, существование таких систем в Т-клетках-хелперах и в макрофагах объяснимо, оно предполагает наличие у них защиты от ретровирусов. Но от каких? Белок, кодируемый локусом генов APOBEC 3G, «работает» только с ВИЧ, утратившими ген белка Vif (фактор инфекционности вируса), и не влияет на продукцию вируса с полноценным Vif (S. Wahl et al., 2006; B.G. Cullen, 2006). Следовательно, его основная функция заключается в устранении не всех ВИЧ, а только тех, которые не могут распространяться по популяциям человека. А TRIM5-альфа «отсекает» от ядра клетки человека не ВИЧ-1, а «чужие» для него ретровирусы, например SIV и ВИЧ-2 (Li X. et al., 2007; S.M. Kaiser et al., 2007).
Обе системы можно рассматривать не столько как антиретровирусные, сколько как фильтры, отделяющие «свои» ретровирусы (т.е. ВИЧ-1 и, возможно, другие, еще нам неизвестные) от каких-то других, присутствие которых в геноме современного вида человека «не предусмотрено» эволюционным процессом задолго до его появления.
Если в сравнительном аспекте рассматривать инфекционные процессы, вызванные ВИЧ, и инфекционные процессы, вызванные ВНО и почему-то столь показательные для разработчиков ВИЧ-вакцин, то мы обнаружим, что они на любом этапе противоположны друг другу 5.
Почему так работают антиретровирусные системы человека? Причина, скорее всего, та же, что заставляет иммунную систему человека участвовать в размножении и распространении ВИЧ — эти системы созданы самими ретровирусами и предполагают существование каких-то механизмов конкуренции между самими ретровирусами за геном хозяина. Остатки ретровирусов с повторяющимися элементами (главным образом LTR из ERV класса I) в сумме составляют до 19 % полного локуса APOBEC3 человека. Наиболее интенсивно они представлены в регионах, фланкирующих APOBEC3G и APOBEC3H (S.G. Conticello et al., 2005).
Эволюционная история TRIM5-альфа изучена лучше, чем APOBEC3. По данным S.M. Kaiser et al. (2007), современный вариант этого белка унаследован человекам от какого его эволюционного предка, подвергшегося селективному давлению со стороны ретровируса PtERV1 не позже 3–4 млн лет назад и представлявшего угрозу для австралопитеков. Эпизоотии, вызванные ретровирусом PtERV1, привели к отбору человекообразных приматов, носителей гена белка TRIM5-альфа, у которого в 332-й позиции находилась аминокислота аргинин. У более ранних предков человекообразных в этой позиции находилась аминокислота глютамин. Это предковое состояние сохранилось у гиббонов, орангутанов и горилл. Поэтому TRIM5-альфа человека эффективен против PtERV1, но бессилен против ВИЧ. В геномах шимпанзе и гориллы сохранились остатки провируса PtERV1. В геноме человека эндогенного варианта этого ретровируса нет. Возможно, селективное давление ВИЧ на популяции людей в отдаленной перспективе и приведет к отбору каких-то человеческих линий с TRIM5-альфа предкового типа, как это предполагают S.M. Kaiser et al. (2007), но цену, которую заплатит наш вид за такую «эволюцию», трудно даже представить.
Инфекционный процесс, вызванный ВНО,— это процесс диссеминации вируса фагоцитирующими клетками по органам и тканям, сопровождающийся выбросом молекул межклеточного общения (хемокинов и лимфокинов) в количествах, значительно превышающих физиологическую норму и, соответственно, проявляющийся развитием патологических реакций (т.е. болезни). Но его продолжительность лимитируется Т- и В-клеточными составляющими иммунной системы хозяина. При рассмотрении инфекционного процесса данного типа речь идет не о конкретном количестве суток, в течение которых начинается и прекращается инфекционный процесс, а о способности клеточной и гуморальной иммунной системы его контролировать. Для инфекционной болезни, развивающейся вследствие инфекционного процесса, лимитируемого клеточной и гуморальной иммунной системой, характерны следующие периоды: инкубационный, продромальный, нарастания симптомов, разгара болезни, угасания клинических проявлений болезни, выздоровления (реконвалесценции) с формированием стерильного иммунитета. Поэтому такие инфекционные процессы я предлагаю назвать циклическими инфекционными процессами. Как правило, они представляют собой монопроцессы, т.е. вызываются одним микроорганизмом. Эпидемии, вызываемые микроорганизмами, использующими такую стратегию (стратегия первого типа), обычно прекращаются из-за формирования иммунной прослойки среди населения, угасания активности природного очага или в результате противоэпидемических мероприятий, направленных на разрыв эпидемической цепи. Они и есть те эпидемии, на борьбу с которыми направлены современные противоэпидемические мероприятия (рис. 7).
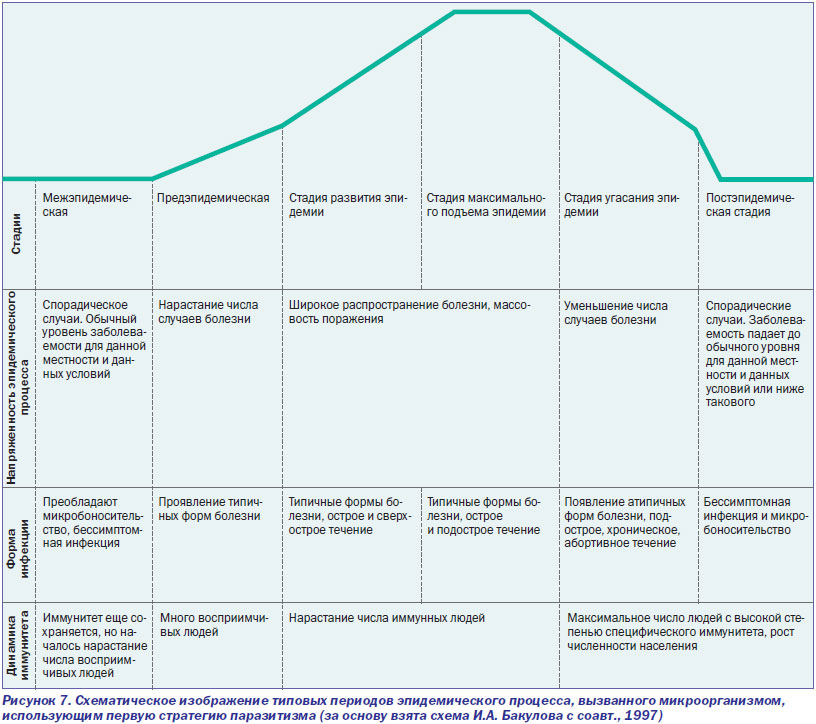
Ответы иммунной системы человека на ВНО и ВИЧ эффективны, но прямо противоположны по содержанию. В отличие от ВНО, пролиферация ВИЧ не контролируется Т- и В-клеточными составляющими иммунной системы. Наоборот, иммунная система помогает вирусу расширить свой ареал за счет фагоцитирующих клеток. Вызываемый таким паразитом инфекционный процесс не блокируется Т- и В-клеточными составляющими иммунной системы. Поэтому он не носит циклический характер, не предполагает периода угасания клинических проявлений болезни и выздоровления больного (реконвалесценции). Передача паразита между хозяевами происходит растянутым во времени, но всегда реализуемым путем — половым, без которого вид не может размножаться. Такая стратегия (стратегия второго типа) дает преимущества паразиту среди особей малочисленных популяций хозяев, обитающих на обширных территориях (рис. 8).
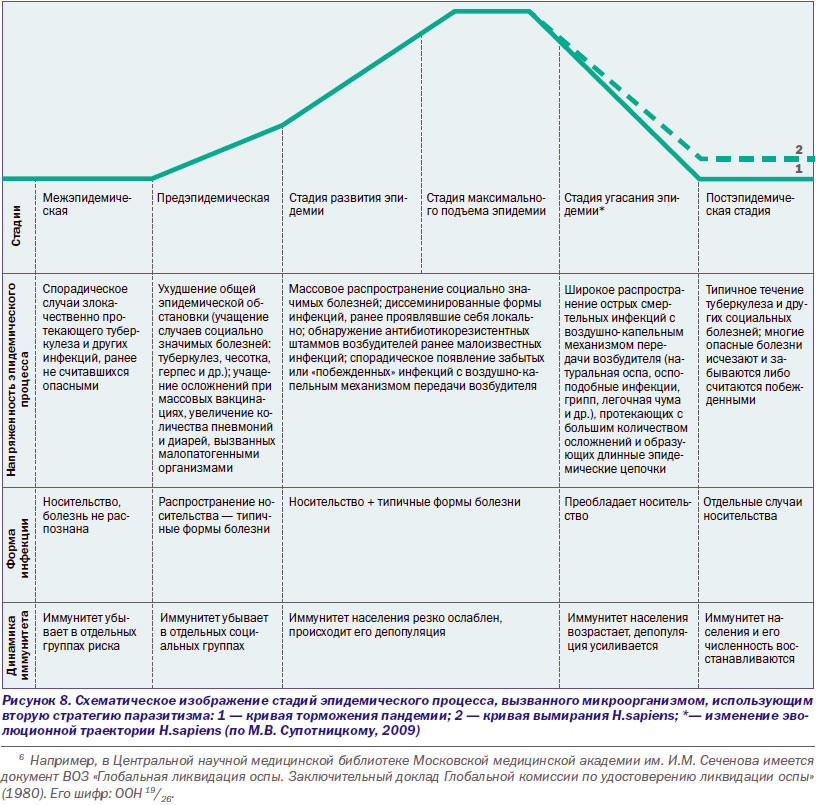
По мере развития ВИЧ-инфекции фагоцитирующие клетки, утратившие контроль со стороны Т- и В-клеточных составляющих иммунной системы, начинают играть в инфекционном процессе ту же роль «мусорщиков», которую они играли у первых многоклеточных животных, что проявляется множеством вялых инфекционных процессов, называемых СПИД-ассоциируемыми. Взаимодействие вызывающих их возбудителей между собой, с ВИЧ и клетками иммунной системы, носит специфический характер, прослеживаемый по крайней мере на надклеточном, клеточном и генетическом уровнях.
Как пример надклеточного специфического взаимодействия можно привести участие белка SP-A в развитии туберкулеза. Присутствующий в бронхоальвеолярной жидкости ВИЧ-инфицированных людей, усиливает прикрепление M.tuberculosis к альвеолярным макрофагам (J.P. Downing et al., 1995).
Клеточное специфическое взаимодействиеиллюстрируется следующими примерами: CDKN1A/p21 — классический ингибитор G1-фазы клеточного цикла индуцируется как ВИЧ, так и микобактериями, но одновременно он является стимулятором жизненного цикла и ВИЧ, и микобактерий в макрофагальной клетке (N. Vazquez et al., 2005); индукция M.avium основного транскрипционного активатора воспалительных цитокинов, NF-кB, ведет к увеличению экспрессии CCR5 и цитокинов, стимулирующих репликацию ВИЧ (S.M. Wahl et al., 2000; S. Wahl et al., 2003; N. Vazquez et al., 2005); экспрессия генов ВИЧ, регулируемая посредством LTR, может быть трансактивирована регуляторными генами многих ДНК-вирусов; они же способны повысить чувствительность к ВИЧ у CD8+ T-клеток и NK-клеток; «переключить» тропность ВИЧ с корецептора CCR5 на корецептор CXCR4 (H.B. Urnovitz, W.H. Murphy, 1996; P. Lusso et al., 2007).
Очень сложны взаимоотношения между эндогенными и экзогенными ретровирусами на уровне генома. Эндогенные ретровирусы и ретроэлементы участвуют в комплементации нарушенных функций экзогенных ретровирусов. Отдельные HERV-K имеют транскрипционно активные открытые рамки считывания и кодируют собственную протеазу, идентичную протеазе ВИЧ. Протеаза HERV-K может комплементировать функцию протеазы ВИЧ у ВИЧ-инфицированных пациентов, подвергнутых лечению ингибиторами протеаз, и тем самым значительно снизить эффективность таких препаратов (M. Padow еt al., 2000). Экзогенные ретровирусы активизируют эндогенные ретровирусы и ретроэлементы. J.J. Goedert et al. (1999) показали усиление экспрессии генов эндогеного ретровируса K10 (HERV-K10) у ВИЧ-инфицированных людей и больных СПИДом и, соответственно, повышение риска развития у них тестикулярного рака (testicular cancer). ВИЧ индуцирует появление вирусных частиц HERV-K (HML-2) в сыворотке человека (R. Contreras-Galindo et al., 2006, 2007).
Следовательно, инфекционный процесс, вызванный ВИЧ, является многокомпонентным и не имеет никаких механизмов самоограничении. Его сложность нарастает по мере ослабления контроля над фагоцитирующими клетками со стороны Т- и В- составляющих иммунной системы. Чтобы понятийно отделить такие процессы от инфекционных процессов, контролируемых Т- и В-составляющими иммунной системы, целесообразно ввести термин «многокомпонентный нециклический инфекционный процесс».
Отдельную проблему составляют «молчащие педиатрические инфекции» (silent pediatric infections). По данным Р. Vаzquez et al. (2006), суть феномена заключается в обнаружении провирусной ДНК ВИЧ в мононуклеарных клетках крови серонегативных детей, родившихся от ВИЧ-инфицированных родителей три и более года назад. Эпидемическая опасность и масштабы распространения феномена неясны. Является ли такой тип течения ВИЧ-инфекции абортивным, или это проявление эндогенизации вируса, или еще что-то другое, станет окончательно ясно только через несколько десятилетий, когда удастся проследить наследование провирусной ДНК ВИЧ у следующих поколений людей.
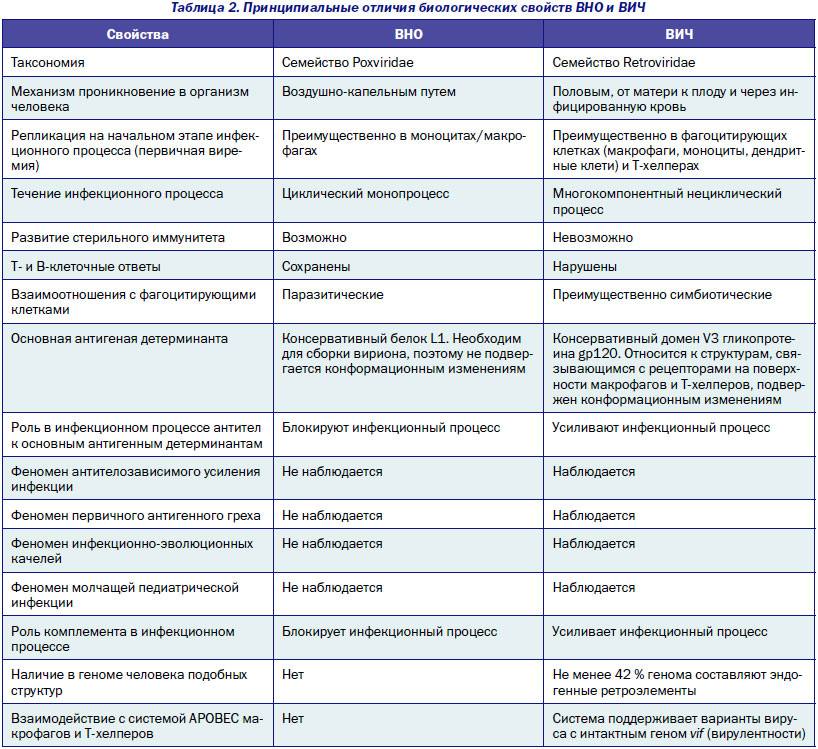
В табл. 2 представлен краткий сравнительный анализ биологических свойств ВНО и ВИЧ, показывающий, что проблема ВИЧ/СПИД-пандемии имеет принципиально иной характер, чем эпидемии периода «до ВИЧ», и читателю целесообразно самому себе задать себе вопрос: «Можем ли мы бороться с ВИЧ/СПИД-пандемией по лекалам, разработанным для борьбы с принципиально иными инфекциями?»
Мы же вновь обратимся к опыту борьбы с натуральной оспой. При всем обилии лжи, окружающей антиВИЧ-мероприятия, самой удручающей является ложь о роли вакцинации в ликвидации натуральной оспы. Удручающей эта ложь является потому, что легко опровергается путем обращения к документам ВОЗ начала 1980-х гг., причем сами эти документы есть в медицинских библиотеках6.
Фактически вся стратегия Программы ликвидации натуральной оспы в глобальном масштабе, провозглашенная ВОЗ в 1959 г., на первом этапе сводилась к массовой вакцинации населения. Но в реальных условиях ликвидации оспы в развивающихся странах использование только массовых прививок оказалось недостаточным. Например, начиная с 1962 г. кампания массовой вакцинации в Индии не привела к сколько-нибудь заметному снижению заболеваемости оспой к 1967 г. Наоборот, через пять лет после начала национальной кампании по ликвидации оспы в 1962 г. число регистрируемых случаев в Индии было выше уровней, отмечавшихся за любой год после 1958 г. В Индонезии на острове Ява, где охват вакцинацией населения превышал 90 %, продолжалась трансмиссия оспы («Глобальная ликвидация…», 1980; С.С. Маренникова, С.Н. Щелкунов, 1998).
На заседании научной группы по ликвидации оспы (октябрь 1967 г.) эксперты рассмотрели ход выполнения как отдельных национальных программ, так и Программы в целом, обращая особое внимание на факторы, оказывающие отрицательное влияние на ее развитие. Наиболее существенным было то, что эта научная группа впервые подчеркнула важное значение эпидемиологического надзора. Эпиднадзор стал основным компонентом второго этапа Программы во всех ее фазах. Проведение систематической вакцинации населения стало рассматриваться как поддерживающая мера. Опыт ряда стран Западной Африки показал, что введение системы активного эпиднадзора позволяет быстро выявлять вспышки оспы и проводить эффективные меры по их ограничению и подавлению с помощью экстренной вакцинации населения этих районов. Новая для программы система оказалась более эффективной для прерывания трансмиссии оспы, чем «поголовная» вакцинация даже в тех случаях, когда было вакцинировано менее половины населения на данной территории. С учетом этих данных Комитет экспертов определил эпиднадзор как краеугольный камень стратегии ликвидации оспы («Глобальная ликвидация…», 1980; Д.А. Хендерсон, 1980; С.С. Маренникова, С.Н. Щелкунов, 1998).
Помимо переоценки роли эпиднадзора, сделанной научной группой в 1967 г. и Комитетом экспертов по ликвидации оспы в 1972 г., чрезвычайно значение имели рекомендация о необходимости введения оценки и контроля каждого компонента Программы и развитие службы регистрации и оповещения о случаях заболевания. Именно контроль за результатами проведенной работы обеспечил реальную базу для принятия правильных решений. Через год, в декабре 1979 г., Глобальная комиссия по удостоверению ликвидации оспы пришла к выводу об успешном завершении Программы в глобальном масшта-бе («Глобальная ликвидация…», 1980; Д.А. Хендерсон, 1980; С.С. Маренникова, С.Н. Щелкунов, 1998).
Теперь давайте посмотрим, что на практике означает перенос этого опыта на борьбу с ВИЧ-пандемией. Возможно ли это в принципе? Сначала придется многократно исследовать сложными иммунологическими и молекулярно-биологическими методами (а не только путем осмотра санитаром кожных покровов) население каждого города или деревни на носительство ВИЧ. Затем выявленных ВИЧ-инфицированных жителей изолировать до конца их жизни (а не на 5–10 сут., как это делали в очагах натуральной оспы в Индии) и только потом оставшееся население многократно вакцинировать ВИЧ-вакциной, если такая вдруг будет создана. Можно ли изъять почти 50 млн ВИЧ-инфицированных человек из эпидемических цепочек? Нет. Тогда зачем пускать пыль в глаза бесконечными разговорами типа: «Вот создадим ВИЧ-вакцину и покончим с ВИЧ, как с натуральной оспой»? Не покончим! Не покончим с ВИЧ/СПИД-пандемией и путем ее тщательного замалчивания, как это имеет место в настоящее время.
Приведенные в статье данные показывают необходимость разработки стратегии борьбы с нециклическими многокомпонентными эпидемическими процессами — это уникальная и не имеющая аналогов в истории медицины задача. Какой будет эта стратегия, пока можно только предполагать. Но нужно прекратить недооценивать и замалчивать проблему ВИЧ/СПИДа, «мир без ВИЧ» остался в нашем победном прошлом.
Список литературы находится в редакции