
Журнал «Здоровье ребенка» 6 (27) 2010
Вернуться к номеру
Природжені компоненти імунітету: Toll-подібні рецептори в нормі і при патології
Авторы: КРЮЧКО Т.О., ТКАЧЕНКО О.Я., ВОВК Ю.О., Вищий державний навчальний заклад України «Українська медична стоматологічна академія», м. Полтава
Рубрики: Педиатрия/Неонатология
Версия для печати
Результати досліджень, проведених за останнє десятиріччя, показали, що імунна система організму має не тільки специфічну глобулінову систему, що характерна для адаптивного імунітету, але й іншу, не менш складну систему розпізнавання чужорідних структур, яка реалізується за допомогою Toll-подібних рецепторів природженої імунної системи. Висвітлено значення Toll-подібного рецептора-4 в реалізації бронхіальної астми у дітей. Розглянуто основні поліморфізми генів TLR4 (Asp299Gly і Thr399Ile) та сигнальні шляхи, які ними запускаються.
Астма, природжена імунна система, Toll-подібний рецептор-4, поліморфізм.
Алергопатологія займає вагоме місце в структурі захворюваності населення багатьох країн світу. З-поміж широкого спектра алергічних захворювань особливу увагу до себе привертає бронхіальна астма (БА), що є серйозною глобальною проблемою сьогодення. Люди всіх вікових груп у багатьох країнах світу зазнають негативного впливу від даного хронічного захворювання дихальних шляхів, яке може бути тяжким, неконтрольованим, перебігати із ускладненнями та в окремих випадках закінчуватись летально.
Починаючи з 1970-х років поширеність астми зросла в більшості країн світу, що пов’язано з урбанізацією та переорієнтацією стилю життя. На сьогодні зареєстровано близько 300 мільйонів хворих на БА і, згідно з прогнозом, до 2025 року цей показник зросте до 400 мільйонів хворих [15]. Крім того, в індустріальному світі БА — найчастіше хронічне захворювання дитячого віку, що прямо пропорційно характеризує стан імунітету дитячого населення, схильність до формування гіперчутливих реакцій, вплив неантропогенних та антропогенних чинників. Створюючи навколо себе штучне місце існування і перемагаючи багато підступних недуг, людство зіткнулося з новим серйозним викликом з боку власної природи. Той факт, що саме в західних країнах частота БА є найбільш високою, спонукав сформулювати так звану гігієнічну гіпотезу, запропоновану ще в 1989 році, яка засвідчила взаємозв’язок між експозицією мікроорганізмів та захистом від алергії.
Важливу роль у первинному захисті організму людини від патогенів відіграє природжена імунна система. Тому протягом останнього десятиріччя увага науковців сконцентрована на виявленні порушень у функціонуванні даної системи як основи для реалізації алергопатології, та зокрема БА. Загальновідомим є те, що взаємодія макроорганізму із навколишнім середовищем реалізується шляхом нормального функціонування дендритних клітин — основних антигенпрезентуючих структур організму людини. Перехід від неактивної до активної форми дендритних клітин є ключовим моментом в активації адаптивної імунної системи. На думку багатьох учених, такий механізм створює стійкий сигнал природженої імунної відповіді, який являє собою стародавню форму імунної системи з подвійним значенням для макроорганізму: забезпечує безпосередній захисний механізм проти мікроорганізмів та виконує роль пам’яті, впливаючи, таким чином, на дозрівання адаптивної імунної системи [2].
Усвідомлення важливої ролі природженого імунітету прийшло разом з ідентифікацією патерн-розпізнавальних рецепторів (ПРР, PRR, pattern recognition receptors), відомих у сучасній науковій літературі як Toll-подібні рецептори (TLR) [14]. Вони належать до групи сигнальних патерн-розпізнавальних рецепторів і розглядаються дослідниками в ролі ключових рецепторів природженого імунітету, оскільки TLRs забезпечують молекулярну ідентифікацію патогену із подальшим включенням важливих компонентів спадкового імунітету та відіграють вирішальну роль у ранньому захисті організму від чужорідних структур [13]. Функціонуючи на зовнішній мембрані моноцитів, макрофагів, нейтрофілів, еозинофілів та тучних клітин макроорганізму, TLRs мають характерну властивість розпізнавання молекулярних структур мікроорганізмів, які отримали назву PAMP (pathogen-associated molecular patterns). Іще в 1989 році, незважаючи на особливості будови різних видів мікроорганізмів, Карл Дженвей сформулював «гіпотезу розпізнавання образів», суть якої полягала в тому, що всі патогенні мікроорганізми мають еволюційно консервативні молекулярні структури, які асоційовані з патогеном і яких немає в багатоклітинних організмах. Підтвердження такої гіпотези прийшло після відкриття у плодової мухи дрозофіли Toll-рецептора, який відповідав за дорзовентральну поляризацію в ембріональному розвитку мухи. Після детального його вивчення виявлено, що в мутаційній популяції дорослих мух із відсутністю білка Toll спостерігалась різко підвищена схильність до грибкової інфекції в результаті зниженої продукції протигрибкового фактору дорзоміцину, що підтвердило важливість ПРР у функціонуванні природженого імунітету. В 1997 році в лабораторії К. Дженвея на моноцитах людини вперше виявлено рецептор, гомологічний Toll-рецептору дрозофіли, тому він і отримав назву Toll-подібний рецептор. Методом позиційного клонування гену, який відповідальний за розпізнавання ліпополісахариду, була підтверджена роль TLR в імунному захисті організму. Надалі йому був присвоєний номер 4 (TLR4). Фактично до початку ХХІ ст. зібрана величезна база даних, яка підтверджує: реакція природженого імунітету на вплив патогенних мікроорганізмів — це специфічна реакція, що здійснюється через патерн-розпізнавальні рецептори в клітинах організму.
Відкриття лігандів TLR відбувалось паралельно з відкриттям самих рецепторів: у першу чергу вивчались консервативні молекулярні структури патогенних мікроорганізмів, надалі була доведена специфічність їх взаємодії з Toll-подібні рецепторами (рис. 1). На сьогодні визначена лігандна специфікація для всіх 10 ПРР, наприклад, TLR2 зв’язується з ліпопротеїдами грампозитивних бактерій; TLR3 — із вірусною двоспіральною РНК; TLR4 — із ліпополісахаридом грамнегативних бактерій; TLR5 зв’язується з флагеліном джгутикових бактерій; TLR6 — із ліпотейхоєвою кислотою (компонент грампозитивних бактерій) та зимозаном, який виділяється дріжджовими мікроорганізмами; TLR7 та TLR8 зв’язуються із вірусною односпіральною РНК; TLR9 взаємодіє з CpG-збагаченою ділянкою ДНК бактерій або вірусів.
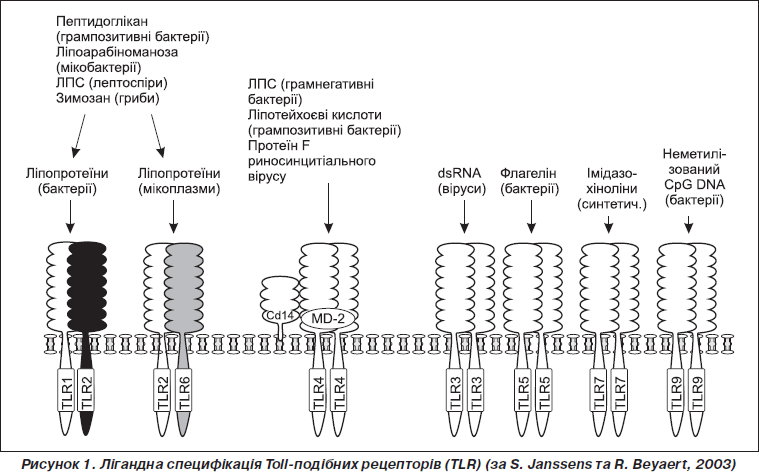
Наведені приклади лігандів досить довго вважались єдиними для кожного типу рецепторів сімейства TLR, проте поступово відкривалися нові види лігандів, розширюючи, таким чином, можливості сімейства Toll. Під час становлення імунології TLR виявилось, що їхніми лігандами можуть бути не тільки компоненти мікроорганізмів, але і речовини іншого походження, наприклад молекулярні структури рослин (TLR4 зв’язується з таксолом), алергенів (наприклад, TLR4 активується екстрактами домашнього пилу), ендогенні сполуки організму (наприклад, білки теплового шоку (HSP) [1]. Таким чином, специфічними лігандами для TLRs є молекулярні структури не тільки мікроорганізмів, але й широке коло різноманітних молекул макроорганізму, рослин та синтетичних речовин.
Іншим, не менш важливим відкриттям став доказ того, що активація ПРР та сигнальні шляхи, які ними запускаються, є стародавньою еволюційно-консервативною системою, що здатна не тільки розпізнавати патогени, але й ініціювати комплекс захисних механізмів організму. У результаті специфічного контакту з лігандом запускається синтез цитокінів за рахунок активації транскрипції генів у мРНК, які відповідальні за їх продукцію. Паралельно досліджувались сигнальні шляхи від активованого ПРР до ядра клітини. На сьогодні ці складні багатокомпонентні шляхи вивчені практично для всіх відомих патерн-розпізнавальних рецепторів.
Слід зазначити, що шляхи активації клітини, ініційовані різноманітними зовнішніми та внутрішніми факторами, здійснюються за єдиним молекулярним принципом [4, 6]. Увесь сигнальний шлях активації TLR складний та багатокомпонентний, але кінцевим етапом каскаду реакцій протеїнкіназ є активація транскрипційних факторів, що знаходяться в цитолізі клітини в заблокованому (неактивному) стані. Натепер відомо декілька груп транскрипційних факторів, але найбільш вивченим є нуклеарний фактор kB (NF-kB). Значення протеїнкіназ полягає в тому, що після звільнення від блокатора нуклеарний фактор переходить в ядро клітини, де зв’язується з промоторними ділянками індуцибельного гену, що, в свою чергу, призводить до його активації та запуску синтезу молекул, які він кодує шляхом утворення специфічних матричних РНК, активації запальних реакцій, у тому числі генів цитокінів.
З-поміж усіх ПРР TLR4 є унікальним, оскільки може взаємодіяти з MyD88 або TIRAP, індукуючи синтез протизапальних цитокінів, або з TICAM-1 і TICAM-2, що приводить до синтезу інтерферонів. Адаптерні білки зв’язуються зі специфічними ферментами-кіназами, що значно підсилюють сигнал та призводять до індукції певних генів, які визначають запальну відповідь клітини [1, 8]. Toll-подібний рецептор-4 є одним із найсильніших клітинних модуляторів, що виконує роль основного захисного механізму від атопічних захворювань, та зокрема БА. Тому точкові мутації на рівні ДНК, які кодують структуру рецептора, можуть призвести до порушення його нормального функціонування. Слід зазначити, що найчастішою причиною відмінностей у структурі генів є так званий поліморфізм одиничних нуклеотидів (SNP-single-nucleotide polymorphism), який за рахунок формування специфічних алелей генів посилено впливає на особливості розвитку захисних реакцій та схильність до низки захворювань. Поліморфізм генів передбачає, що з одного і того ж гена може бути скопійовано декілька варіантів, які структурно відрізняються від копії одного і того ж білка, із них частина скопійованих варіантів або взагалі неактивна, або може мати протилежну функцію. Основними поліморфізмами одиничних нуклеотидів TLR4 є Asp299Gly та Thr399Ile, які тісно пов’язані з розвитком атопічних захворювань. Крім того, існують результати, які засвідчують, що TLR4 (Asp299Gly) поліморфізм підвищує частоту інфекцій, хронічних запальних процесів у дихальних шляхах [3, 4]. Секвестрація людського TLR4 дала можливість повністю визначити можливі варіації негомологічних поліморфізмів в третьому екзоні, який відповідальний за маркірування лейцинового домену. Але, незважаючи на значні коливання варіацій TLR4 та LRR, частота таких поліморфізмів у людській популяції є дуже низькою (менше 1 %). Виняток становлять два негомологічні поліморфізми (SNPs), частота виявлення яких більше 5 %. Це так зване A/G-заміщення, тобто причиною даного поліморфізму є заміна аспарагінової амінокислоти на гліцинову в положенні Asp299Gly (rs4986790), а також С/Т-заміщення, тобто заміщення треоніну на ізолейцин у положенні Thr399Ile (rs4986791). Уперше такі два поліморфізми (Asp299Gly або Thr399Ile) у людини виявив N.C. Arbour і охарактеризував їх як зниження відповіді імунної системи на вдихання LPS. Відкриття зниженої реактивності на вплив бактеріального ліпополісахариду у людей із генетичними варіаціями TLR4 (Asp299Gly та Thr399Ile) спонукало до проведення багатьох подібних генетичних досліджень [5].
Результати таких досліджень призвели до суперечливих думок з приводу ролі поліморфізму Asp299Gly та його впливу на сприйнятливість до грамнегативних бактеріальних інфекцій [16]. Під час усвідомлення молекулярних механізмів природженого імунітету з’явилась одна з теорій, що змогла пояснити таку відмінність в отриманих результатах. Суть її полягала в тому, що всі проведені дослідження поліморфізмів Asp299Gly та Thr399Ile проводились окремо один від одного, як самостійні форми і зовсім не враховувався той факт, що два поліморфізми можуть поєднуватися в одної особи (Asp299Gly/Thr399Ile). Таке поєднання передбачає виділення чотирьох гаплотипів, представлених у людській популяції, а саме: wt/wt, Asp299Gly/wt, Thr399Ile/wt та Asp299Gly/Thr399Ile. Результати цитокінового профілю після стимуляції ліпополісахаридом засвідчили, що лише Asp299Gly-гаплотип має фенотипові зміни, які виявляються переважно у підвищенні прозапального TNF-a [7]. Цікаво, що LPS-індукована імунна відповідь при Asp299Gly/Thr399Ile-гаплотипі нічим не відрізняється від цитокінової відповіді дикого типу TLR4Asp299Gly/Thr399Ile через те, що окремий гаплотип Thr399Ile зустрічається дуже рідко і тому його прояви на сьогодні залишаються невідомими. Ось чому гаплотип TLR4 змінює цитокінсинтезуючу відповідь за впливу бактеріального ліпополісахариду, але зміна реактивності на вплив грамнегативних інфекцій лежить лише в основі TLR4 Asp299Gly-мутації. Популяційні дослідження також висвітлили специфіку географічного розподілу TLR4-гаплотипу [17]. Сьогодні в африканській популяції Asp299Gly-гаплотип зустрічається в 10–20 разів частіше, ніж на інших континентах; утім у європеоїдній популяції спостерігається майже повна відсутність даного гаплотипу та висока частота виявлення Asp299Gly/Thr399Ile-гаплотипу. В азіатської популяції взагалі були відсутні всі три Asp299Gly-, Thr399Ile- та Asp299Gly/Thr399Ile-гаплотипи.
Оскільки в основі даного поліморфізму лежить генетично обумовлене порушення структури TLR4, логічно передбачити, що подібні зміни на рівні ДНК та обумовлені ними нозології можуть передаватися і наступним поколінням. Наприклад, дослідження, проведене у великій когорті волонтерів, засвідчило асоціацію між наявністю поліморфізму TLR4 (Asp299Gly) та розвитком бронхіальної астми в декількох поколіннях одного родоводу. Звичайно, частота подібних SNP зустрічається рідко. За даними різних досліджень, вона не перевищує 5 % загальної кількості хворих на БА [9]. Проте дійсно генетична обумовленість БА з виявленим поліморфізмом TLR4 (Asp299Gly) беззаперечна.
На сьогодні особливої актуальності набуває вивчення поліморфізму генів регуляторних молекул запалення. Знання їхньої ролі в патогенезі багатьох хвороб поряд із досягненнями сучасної геноміки дозволяє прогнозувати ризик розвитку патології або тяжкість її перебігу та дібрати специфічну терапію або зробити відповідну корекцію у схемі лікувально-профілактичних заходів. Оскільки РАМР бактерій, вірусів та грибків є сильними активаторами природженої імунної відповіді, то на їх основі створюються високоефективні імунотропні лікарські препарати. За напрямком дії ці препарати можуть бути двох типів: агоністи Toll-подібних рецепторів — імуностимулюючі препарати та антагоністи — імуносупресанти, спрямовані на корекцію хронічних імунозапальних процесів. На даному етапі розробка таких препаратів ведеться за кордоном. Усі вони знаходяться лише на етапі клінічних випробувань. Наприклад, мінімально біологічно активні фрагменти, виділені з ліпополісахариду — монофосфорил-ліпід А (агоніст TLR4) вже використовують як ад’юванти, що входять до складу вакцин з метою посилення їх імуногенних властивостей.
На завершення слід зазначити, що активне дослідження природженого імунітету протягом останніх 10 років принесло значні результати не тільки з точки зору фундаментальних уявлень про організацію протиінфекційного імунітету, але й з точки зору практичної медицини. Фактори ризику та механізми реалізації спадково обумовленої патології у людини потребують всебічного вивчення, оскільки ефективна антенатальна діагностика генетично обумовленої атопії передбачає досконалу профілактику шляхом формування груп ризику щодо відтворення даної патології ще до настання вагітності.
1. Лебедев К.А., Понякина И.Д. Иммунология образраспознающих рецепторов. — Москва: ЛИБРОКОМ. — 2008. — 253 с.
2. Семенов Б.Ф., Зверев В.В. Концепция создания быстрой иммунологической защиты от патогенов // Журнал микробиологии, эпидемиологии и иммунобиологии. — 2007. — № 4. — С. 93-100.
3. Толстопятова А.М., Семенов Б.Ф., Зверев В.В. Роль рецепторов врожденного иммунитета в развитии инфекционной патологи у новорожденных детей // Педиатрия. — 2009. — Т. 87, № 1. — С. 115-120.
4. Потехина Е.С., Надеждина Е.С. Митогенактивируемые протеинкиназные каскады и участие в них Set20 подобных протеинкиназ // Успехи современной химии. — 2002. — Т. 2, № 1. — С. 235-255.
5. Arbour N.C., Lorenz E., Schutte B.C. et al. TLR4 mutations are associated with endotoxin hyporesponsiveness in humans // Nat. Genet. — 2000. — № 25. — Р. 187-191.
6. Bengtsson S.L., Nagy Z., Skare S. Extensive piano practicing has regionally specific effects on white matter development // Nat. Neurosci. — 2005. — Vol. 8, № 9. — P. 1148-1150.
7. Ferwerda B., McCall М.В. TLR4 polymorphisms, infectious diseases, and evolutionary pressure during migration of modern humans // Proc. Natl. Acad. Sci USA. — 2007. — Р. 1034-1037.
8. Horner A.A., Raz E. Do microbes influence the pathogenesis of allergic diseases? Building the case for Toll-like receptor ligands // Curr. Opin. Immunol. — 2003. — 15(6). — 614-9.
9. Kormann M.S.D. Toll-like receptor heterodimer variants influenceTh1/Th2 signaling and protect from the development of childhood asthma. — Hannover, 2008. — Р. 110.
10. Leynaert B., Neukirch C. et al. Does living on a farm during childhood protect against asthma, allergic rhinitis, and atopy in adulthood? // Am. J. Respir. Crit. Care Med. — 2001. — 164(10 Pt1). — Р. 1829-34.
11. Matricardi P.M., Rosmini F. et al. Exposure to foodborne and orofecal microbes versus airborne viruses in relation to atopy and allergic asthma: epidemiological study // BMJ. — 2000. — 320(7232). — Р. 412-7.
12. Martinez F.D., Holt P.G. Role of microbial burden in aetiology of allergy and asthma // Lancet. — 1999. — 354 Suppl 2. — SII12-5.
13. Medzhitov R., Preston-Hurlburt P. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity // Nature. — 1997. — 388(6640). — P. 394-7.
14. Medzhitov R. Toll-like receptors and innate immunity // Nat. Rev. Immunol. — 2001. — Vol. 1(2). — P. 135-45.
15. Rees J. Контроль бронхиальной астмы у взрослых: Переклад з англ. // Здоров’я України. — 2007. — № 2. — С. 33-34.
16. Schroder N.W., Schumann R.R. Single nucleotide polymorphisms of Toll-like receptors and susceptibility to infectious disease // Lancet Infect. Dis. — 2005. — № 5. — Р. 156-164.
17. Smirnova I., Hamblin М.Т., McBride С., Beutler В. Excess of rare amino acid polymorphisms in the Toll-like receptor 4 in humans // Genetics. — 2001. — № 158. — Р. 1657-1664.