
Международный эндокринологический журнал 3 (35) 2011
Вернуться к номеру
Ультраструктурная характеристика тиреоцитов при гипо- и гиперфункции щитовидной железы
Авторы: Юлдашева Ф.З., Юлдашев А.Ю., Исмаилов С.И., Рашитов М.М., Республиканский специализированный научно-практический медицинский центр эндокринологии МЗ РУз, г. Ташкент, Узбекистан
Рубрики: Эндокринология
Версия для печати
На модели состояния дефицита йода (I гр.), эутиреоза (II гр.), избыточного содержания (III гр.) изучена ультраструктура щитовидной железы. Снижение йода в рационе и гипофункция щитовидной железы морфологически характеризуются уменьшением массы органа, высоты и площади тиреоцитов, гипофункцией цитоплазматических структур с уменьшением синтеза секреторного продукта. Высокое содержание йода в рационе вызывает увеличение массы и функции щитовидной железы, высоты и площади тиреоцитов, профилей и объема цитоплазматических структур, увеличение синтеза и секреции секреторных гранул. На основании полученных результатов установлена тесная зависимость между содержанием йода в рационе, массой железы и структурой тиреоцитов.
Щитовидная железа, йод, фолликулы, тиреоциты.
Введение
Щитовидной железе (ЩЖ) принадлежит важное место в регуляции адаптивных реакций организма. Благодаря адекватному изменению ее структуры и функции во всем организме и его системах наблюдаются взаимосвязанные и взаимообусловленные приспособительные процессы. В связи с этим представляет интерес изучение состояния тиреоцитов ЩЖ при избыточном и недостаточном введении йода. Настоящая работа является фрагментом комплексного исследования по изучению механизмов регуляции адаптивных реакций регуляторных систем, органов на различные воздействия.
Цель работы: изучение ультраструктуры щитовидной железы при избыточном или недостаточном содержании йода в организме.
Материалы и методы исследования
Белым беспородным крысам массой 100–130 г, у которых по специально разработанной модели добивались состояния дефицита йода (I гр.), эутиреоза (II гр.), йод из расчета 200 мкг/кг веса крыс введен с питьевой водой [1, 2], избыточного содержания (III гр.) — йод из расчета 1000 мкг/кг. Через 1 месяц наркотизированные животные забивались путем декапитации. У них измеряли массу тела, ЩЖ. Гистологически путем свето- и электронной микроскопии изучены тиреоциты фолликулов ЩЖ. ЩЖ фиксировалась в 2,5% забуференном растворе глютаральдегида (20 мин) с последующей дофиксацией в 1% растворе осмиевой кислоты. После промывки в растворе буфера и проводки через спирты возрастающей концентрации ткань железы залита в аралдит. Срезы получены на ультрамикротоме LKB-4800. После контрастирования в уранилацетате и цитрате свинца ультратонкие срезы просмотрены в микроскопе JEM-100S. На полутонких окрашенных основным фуксином и метиленовым синим срезах железы в каждом случае определяли средний диаметр (Д) 50 фолликулов путем их измерения в двух перпендикулярных направлениях расстояния между апикальными поверхностями тиреоцитов. Высота (Н) тиреоцитов измерялась от базальной части клетки до апикальной. Отношение Н клетки к Д фолликула служило характеристикой функциональной активности железы. На каждый случай эксперимента (I–III гр.) использована ЩЖ не менее 6 животных.
Площадь ядер и цитоплазмы не менее 100 тиреоцитов у каждого отдельного животного определялась с помощью полуавтоматического анализатора изображений «Интеграл-2М».
Статистическая обработка количественных результатов осуществлялась с помощью компьютерных программ Microsoft Excel и Statistica 6.
Результаты исследования и их обсуждение
У крыс II группы 100 г массы тела соответствует в среднем 15,24 ± 0,88 мг ЩЖ. Паренхима железы снаружи покрыта соединительнотканной капсулой. От нее в паренхиму железы отходят перегородки, где располагаются нервы, сосуды, кровеносные и лимфатические сосуды. В каждой дольке железы различаются овальные или округлые фолликулы. На одной общей базальной мембране, ограничивающей каждый фолликул, одним слоем располагаются кубической формы тиреоциты (рис. 1). Полость фолликула заполнена эозинофильным коллоидом. При диаметре фолликула 84,6 ± 4,5 усл.ед. высота тиреоцита составляет 68,2 ± 0,6 усл.ед. Отношение Н тиреоцитов к Д фолликула как показатель функциональной активности железы равно 1,24.
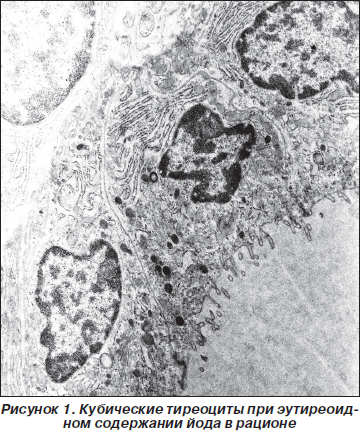
Ядра тиреоцитов располагаются в базальной их части, содержат 1–2 электронно-плотных ядрышка. Нуклеоплазма богата эухроматином, гиперхроматин узкой полоской обнаруживается под нуклеолеммой. На свободной поверхности тиреоцитов выявляются до 10–12 микроворсинок длиной 0,2–0,4 мкм. Боковая плазмолемма образует на уровне границы с полостью фолликула соединительнотканный комплекс, герметизирующий межэпителиальное пространство. Ниже его различаются десмосомы и интердигитации. Базальная плазмолемма тиреоцитов формирует различные по числу и выраженности складки, вдающиеся в цитоплазму, благодаря чему ее протяженность увеличивается. Профили шероховатого эндоплазматического ретикулума (ШЭР) занимают значительную часть цитоплазмы и располагаются параллельно друг другу в базальной части и вокруг ядра, вдоль латеральной поверхности клетки. Если образуемые умеренно расширенные цистерны располагаются под ядром, то уплощенные — вдоль боковой поверхности ядра. Комплекс Гольджи, состоящий из цистерны, везикул и вакуолей, располагается над верхним полюсом ядра, занимает значительную зону. Округлые, овальные, удлиненные митохондрии выявляются повсеместно: между цистернами ШЭР, вокруг структур комплекса Гольджи. Кристы ориентированы поперечно, в умеренном количестве, находятся в матриксе средней плотности. Свободные рибосомы и полисомы в большом количестве равномерно рассеяны по всей цитоплазме.
Секреторные гранулы умеренной и значительной плотности, в диаметре 400–500 А, локализуются в апикальной части тиреоцитов. Кроме них в этой части клетки имеются также капли коллоида с диаметром 400–1500 А, низкой, средней и высокой электронной плотности.
Наряду с тиреоцитами в каждом фолликуле различаются 2 типа парафолликулярных клеток, которые своей поверхностью не контактируют с коллоидом. Если один из них аналогичен по ультраструктуре тиреоцитам, отличается отсутствием признаков секреторной активности, гранул в цитоплазме, то второй тип — С-клетки располагаются поодиночке, по 2–3, имеют параллельно расположенные 2–3 профиля ШЭР, образующих цистерны и канальцы. Комплекс Гольджи в них хорошо развит, локализован около ядра. Митохондрии овальные, округлые, с матриксом умеренной плотности. Кристы ориентированы, в умеренном количестве. Характерным для С-клеток является наличие многочисленных мелких электронно-плотных секреторных гранул с диаметром 100–140 А. Они накапливаются по периферии цитоплазмы клеток, особенно в участках, контактирующих с кровеносными капиллярами. Между фолликулами в строме железы часто встречаются кровеносные капилляры, выстланные уплощенным фенестрированным эндотелием со значительным числом пор и эндоцитозных везикул вдоль люминальной и базальной поверхности. Часто базальные мембраны тиреоцитов и эндотелия близко прилегают друг к другу, отделены тонкой полоской интерстиция.
При длительном (не менее 1 мес.) пребывании животных на диете с низким содержанием йода (I гр.) масса ЩЖ снижается до 11,40 ± 0,65 мг на 100 г массы тела крыс и составляет в среднем 75 % от таковой животных II гр. Значительную часть железы образуют фолликулы крупных размеров, которые выстланы низкокубическими тиреоцитами. Диаметр фолликула увеличивается до 190 ± 11 усл.ед. Это в среднем на 185 % больше, чем у крыс II группы. Высота тиреоцитов снижается до 35,4 ± 0,9 усл.ед., что в среднем на 50 % меньше исходной их высоты. Диаметр фолликулов становится равным 1,00 : 5,37 (у II гр. 1,00 : 1,24).
Если измерить площадь цитоплазмы и ядра, выявляется их уменьшение через 1 месяц после введения гипойодной диеты. Ядра имеют площадь 3,83 ± 0,30 (во II гр. 13,55 ± 0,60). Площадь цитоплазмы тиреоцитов уменьшается до 15,70 ± 0,70 (II гр. — 28,3 ± 0,9). Уменьшение площади цитоплазмы до 55 % (т.е. на 45 %) коррелирует с изменением площади ядра. Ядерно-цитоплазматическое отношение равно 1 : 60 (во II группе 1,00 : 2,05).
Коллоид, который находится в центре каждого фолликула, становится более эозинофильным. Между фолликулами наблюдаются островки скопления парафолликулярных клеток.
Микроскопически через 1 месяц после пребывания на гипойодной диете тиреоциты низкокубические, коллоид более электронно-плотный, чем у эутиреоидных животных II группы (рис. 2). На апикальной поверхности тиреоцитов выпячивания апикальной плазмолеммы единичные, короткие. Латеральная плазмолемма сложных и глубоких интердигитаций не образует. Базальная плазмолемма сглажена, складок не имеет. Цитоплазматические структуры тиреоцитов немногочисленны: профили цитоплазматической сети уплощены или имеют вид везикул и единичных вакуолей. Небольшое число мелких митохондрий выявляется повсеместно; они имеют плотный матрикс и редкие кристы. Комплекс Гольджи находится над ядром и состоит из небольшого числа пузырьков. Полисомы, свободные рибосомы не-многочисленны. Секреторные гранулы в тиреоцитах единичны или не выявляются, имеют низкую плотность. Часто в цитоплазме клеток обнаруживаются округлые липидные включения.
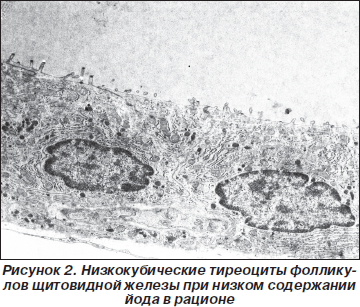
Располагающиеся между фолликулами капилляры выстланы уплощенным эндотелием, однако в них существенно меньше эндоцитозных везикул и пор. В интерстиции увеличивается количество коллагеновых волокон.
Таким образом, через один месяц после гипойодной диеты наблюдаются уменьшение массы ЩЖ, отчетливые признаки снижения функциональной активности тиреоцитов, увеличения диаметра фолликулов.
Если крысы находятся на диете с высоким содержанием йода (III гр.), то масса ЩЖ становится равной 17,01 ± 0,90 мг на 100 г веса, что в среднем на 11 % больше, чем у контрольных животных. Морфологически в паренхиме ЩЖ наряду с крупными фолликулами по периферии железы появляется много мелких в центральной ее части. Стенка фолликулов выстлана призматическими клетками. Коллоид в центре фолликулов относительно менее плотный, чем при сниженной функции железы. Над апикальной частью тиреоцитов коллоид просветлен или вакуолизирован.
При морфометрии отмечается увеличение тиреоцитов до 72,4 ± 0,9 (во II гр. 68,2 ± 0,6; р < 0,05); диаметр фолликула почти не меняется (84,6 ± 4,5 во II группе и 89,1 ± 4,8 в III группе; р > 0,05). Соотношение высоты тиреоцитов и диаметра фолликулов становится равным 1,00 : 1,23 (в контроле 1,00 : 1,24; р > 0,05). Полученные результаты свидетельствуют о соразмерном увеличении высоты тиреоцитов и диаметра фолликулов при увеличении йода в рационе опытных животных.
При измерении площади ядер отмечается сохранение ее параметров, как у эутиреоидных животных II группы (13,40 ± 0,80; р > 0,05). Площадь цитоплазмы становится в среднем на 20 % больше, чем у животных II группы (33,4 ± 1,4 против 28,3 ± 0,9 в контроле; р < 0,05). В результате соотношение равно 1,00 : 2,49 (в контроле 1,00 : 2,05), т.е. увеличение площади цитоплазмы клеток вызывает уменьшение ядерно-цитоплазматического отношения у опытных животных.
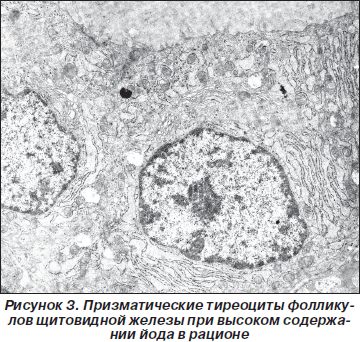
При увеличении содержания йода в рационе в фолликулах выявляются ЩЖ призматические тиреоциты, коллоид внутри умеренной плотности (рис. 3). В апикальной части тиреоциты имеют много удлиненных микроворсинок, куполообразных выпячиваний, базальная плазмолемма образует множество глубоких выпячивающихся в цитоплазму складок тиреоцитов. Капилляры, прилегающие к фолликулам, отделены тонкой прослойкой соединительной ткани, гомогенной пластинкой базальной мембраны. Эндотелий, выстилающий их просвет, уплощен, имеет большое число эндоцитозных везикул на обеих поверхностях. Характерным является наличие в цитоплазме тиреоцитов большого числа расширенных профилей гранулярной цитоплазматической сети вокруг ядра. Митохондрии крупные, удлиненные, с плотным матриксом и поперечно ориентированными кристами. Комплекс Гольджи занимает значительную зону над ядром и состоит из расширенных цистерн, большого числа вакуолей и везикул. В апикальной части тиреоцитов содержится большое количество секреторных гранул, различных по плотности и величине. Ядро располагается в базальной половине клетки, содержит много эухроматина. В этой же части тиреоцитов выявляются коллоидные капли и полиморфные лизосомы.
Выводы
1. Между содержанием йода в рационе, массой щитовидной железы и структурой тиреоцитов имеется тесная зависимость.
2. Снижение йода в рационе и гипофункция щитовидной железы морфологически характеризуются уменьшением массы органа, высоты и площади тиреоцитов, синтеза секреторного продукта, гипофункцией цитоплазматических структур.
3. Высокое содержание йода в рационе вызывает увеличение массы и функции щитовидной железы, высоты и площади тиреоцитов, профилей и объема цитоплазматических структур, увеличение синтеза и секреции секреторных гранул.
1. Туракулов Я.Х., Ташходжаева Т.П., Кулимбетов М.Т. и др. Экспериментальное моделирование гипотиреоза, обусловленного хроническим дефицитом йода в диете // Узбек. биологический журнал. — 1994. — № 1. — С. 12-15.
2. Hotz Ch.S., Fitzpatrick D.W., Trick K.D., L’Abbe M.R. Dietary iodine and selenium interact to affect thyroid hormone metabolism of rats 1–4 // ANZCCART News. — 1993. — № 4.