Журнал «Здоровье ребенка» Том 12, №6, 2017
Терапевтический потенциал бактериофагов и эндолизинов при лечении острых респираторных инфекций, вызванных бактериальными патогенами
Авторы: Абатуров А.Е.(1), Крючко Т.А.(2)
(1) — ГУ «Днепропетровская медицинская академия МЗ Украины», г. Днепр, Украина
(2) — ВГУЗУ «Украинская медицинская стоматологическая академия», г. Полтава, Украина
Рубрики: Педиатрия/Неонатология
Разделы: Справочник специалиста
Резюме
Бактеріофаги являють собою віруси, які за рахунок рецептор/ліганд-взаємодії здатні специфічно зв’язуватися з бактеріальною стінкою і відновлюватися всередині певних мікроорганізмів. Літичні ферменти бактеріофагів ефективно руйнують клітинну стінку бактерій, але на відміну від антибіотиків не впливають на життєдіяльність симбіотичної флори. На сьогодні існує велика кількість наукових матеріалів, що стосуються ефективності антистрептококових, антистафілококових, антиклебсієльозних, антисинегнійних бактеріофагів і ендолізинiв, які здатні викликати загибель антибіотикорезистентних бактерій, у зв’язку з чим їх використовують як бактерицидні лікарські засоби, що є потенційно ефективним терапевтичним методом протимікробної терапії. Високий рівень ефективності бактерицидної дії ендолізинів дозволяє висловити припущення, що майбутні лікарські засоби, створені на молекулярній основі ендолізинів, можуть стати ключовими протимікробними препаратами при лікуванні інфекційних захворювань.
Бактериофаги представляют собой вирусы, которые за счет рецептор/лиганд-взаимодействий способны специфически связываться с бактериальной стенкой и реплицироваться внутри конкретных бактерий. Литические ферменты бактериофагов эффективно разрушают клеточную стенку бактерий, но в отличие от антибиотиков не влияют на жизнедеятельность симбиотической флоры. Сегодня уже собран достаточный иcследовательский опыт эффективности антистрептококковых, антистафилококковых, антиклебсиеллезных, антисинегнойных бактериофагов и эндолизинов, которые способны вызывать гибель антибиотикорезистентных бактерий, в связи с чем их использование в качестве бактерицидных лекарственных средств является потенциально эффективным терапевтическим методом противомикробной терапии. Высокий уровень эффективности бактерицидного действия эндолизинов позволяет предположить, что будущие лекарственные средства, созданные на молекулярной основе эндолизинов, могут стать ключевыми противомикробными препаратами при лечении инвазивных инфекций.
Bacteriophages are viruses that, due to the receptor/ligand interactions, are able to specifically bind to the bacterial wall and replicate within specific bacteria. The lytic enzymes of bacteriophages efficiently destroy the bacterial cell wall, but unlike antibiotics, they do not affect the vital activity of the symbiotic flora. Today, there is sufficient research experience on the efficacy of antistreptococcal, antistaphylococcal, antiklebsiella, antipseudomonal bacteriophages and endolysins that can kill antibiotic-resistant bacteria, and therefore, their use as bactericidal drugs is a potentially effective therapeutic method of antimicrobial therapy. The high level of effectiveness of the bactericidal action of endolysins suggests that future drugs, created on the molecular basis of endolysins, can become key antimicrobial agents in the treatment of invasive infections.
Ключевые слова
гострі респіраторно-вірусні інфекції; діти; бактеріофаги
острые респираторные инфекции; дети; бактериофаги
acute respiratory infections; children; bacteriophages
Введение
Бактериофаги представляют собой вирусы, которые за счет рецептор/лиганд-взаимодействий способны специфически связываться с бактериальной стенкой и реплицироваться внутри конкретных бактерий. Бактериофаги являются наиболее распространенным биологическим объектом в биосфере, представительство которых составляет 1031 организмов, в то время как общее число прокариотических клеток в биосфере составляет около 1030 [7]. Бактериофаги находятся в любой среде, в которой присутствуют бактерии. В водных системах численность популяции бактериофагов составляет 104–108 вирионов на 1 мл, в почве — 109 вирионов на 1 грамм [52]. В настоящее время идентифицировано более 5500 различных бактериофагов [1]. Из идентифицированных фагов большинство принадлежит к хвостатым фагам, которые образуют таксономический порядок: Caudovirales. Данные бактериофаги имеют икосаэдрические головки, содержащие геномы, состоящие из двухцепочечной ДНК. Порядок Caudovirales состоит из трех фаговых семейств: Myoviridae — с жесткими сократительными хвостами, Podoviridae — с короткими, не сокращающимися хвостами и Siphoviridae — с длинными гибкими хвостами. Фаги, принадлежащие к другим семействам, имеют изменчивую морфологию с геномами различного состава нуклеиновых кислот [7]. Бактериофаги, вызывающие гибель бактерий, не индуцируют развитие резистентности бактерий к противомикробным препаратам, недорогие в производстве и безопасны для людей [8]. Бактериофаготерапия была успешно использована для лечения инфекционных кожных заболеваний до открытия антибиотиков [45].
Бактериофаги
В центре фаготерапии Института иммунологии и экспериментальной терапии им. Хиршфельда во Вроцлаве проводят экспериментальную терапию бактериофагами против ряда бактериальных заболеваний, в том числе вызванных Acinetobacter, Burkholderia, Citrobacter, Enterobacter, Enterococcus, Escherichia, Klebsiella, Morganella, Proteus, Pseudomonas, Shigella, Salmonella, Serratia, Staphylococcus и Stenotrophomonas. На основании данных, опубликованных сотрудниками данного центра, положительный результат бактериофаготерапии отмечается у 35–50 % пациентов [24].
В зависимости от вида высвобождения новых вирионов из бактерий бактериофаги делят на литические и лизогенные. Литические бактериофаги способны вызывать деструкцию стенки и у антибиотикорезистентных бактерий. Большинство литических бактериофагов используют две группы протеинов для киллинга инфицированных бактерий: холины и эндолизины. Холины участвуют в процессе активации лизиса клеток-хозяев: они перфорируют цитоплазматическую мембрану бактерии, предоставляют эндолизинам доступ к бактериальному пептидогликану (peptidoglycan — PG), а эндолизины непосредственно вызывают лизис бактерии [21].
Система «холин — лизин» отвечает за терминацию цикла заражения фагом в определенный момент времени. Влияние эндолизинов на клеточную стенку бактерий высоко синхронизировано через механизм, объясненный двухстартовой моделью. Согласно данной модели время бактериального лизиса зависит от соотношения содержаний холина и его антагониста антихолина. Соотношение содержаний холина и антихолина строго регулируется через контроль их экспрессии на трансляционном уровне. Нарушение баланса «холин — антихолин» за счет повышения содержания холина приводит к потере целостности плазматической мембраны, что позволяет эндолизину проникнуть к периплазме и разрушить PG бактерии [46].
Было показано, что литические ферменты бактериофагов эффективно разрушают клеточную стенку целевых грамположительных бактерий и в отличие от антибиотиков не влияют на жизнедеятельность симбиотической флоры. Таким образом, бактерио–фаги представляют собой высокоспецифичные средства, тогда как антибиотики, как правило, обладают широким противомикробным действием. И эта специфичность является существенным недостатком бактериофагальных препаратов. Использование в терапии большинства бактериофагов, как правило, не сопровождается развитием побочных эффектов у макроорганизмов и развитием антибиотикорезистентности у бактерий [13, 41].
Отличия лечения бактериофагальными препаратами от антибактериальной терапии представлены в табл. 1.
Эндолизины
Одновременно с фаготерапией предпринимаются попытки разработать антибактериальные агенты из бактериостатических и бактериолитических белков, кодируемых генами фагов [35]. Так, для лизирования патогенных бактерий предлагается использовать: виролизин, лизин, лизоцим, мурализин, мурамидазу, фагоассоциированные ферменты, фаголизин, фаголизоцим, эндолизины, энзибиотики и другие. В настоящее время в фаготерапии сделан акцент на литические энзимы как возможные целевые противобактериальные прототипы [5, 22, 55].
На молекулярном основании специфических эндолизинов, продуцируемых бактериофагами, создаются новые рекомбинантные лекарственные средства, способные вызвать гибель исключительно причинно-значимых бактериальных агентов [48]. В частности, Vincent A. Fischetti и соавт. [34] продемонстрировали, что очищенный лизин CPL1 способен in vivo и in vitro привести к гибели бактерий большинства пневмококковых серотипов. Авторы предложили название «энзибиотик» для обозначения данных ферментов, участвующих в лизисе стенки бактерий.
В настоящее время наиболее изученными энзимными продуктами бактериофагов являются пептидогликангидролазы (peptidoglycan hydrolases — PGH), или эндолизины. Эндолизины представляют собой ферменты, которые обладают эндопептидазной, амидазной, гликозидазной или трансгликозилазной активностью, вызывающей деградацию PG клеточной стенки бактерии. Действуя в конце цикла репликации фага, эндолизины способствуют высвобождению новых вирионов. Эндолизины, направленные против грамположительных и грам–отрицательных бактерий, структурно отличаются друг от друга, так как стенки разных бактерий существенно отличаются молекулярной архитектурой. Стенка грамположительных бактерий состоит из цитоплазматической мембраны и толстого слоя PG (до 40 слоев), который легко доступен для экзогенных эндолизинов. Эндолизины разрушают слой PG с последующим осмотическим лизисом и гибелью клеток. Стенка грамотрицательных бактерий характеризуется наличием тонкого слоя PG (1–2 слоя), который защищен дополнительной внешней мембраной. Эндолизины, направленные против грамотрицательных бактерий, представляют собой маленькие глобулярные протеины, молекула которых состоит только из одного домена, называемого энзиматически активным доменом (enzymatically active domain — EAD), тогда как молекулы эндолизинов, направленных против грамположительных бактерий, содержат еще один домен — домен связывания клеточной стенки (cell wall binding domain — CBD). Домен CBD вносит свой вклад в литическое действие эндолизина за счет синергизма с EAD, который выполняет каталитическую функцию ферментативного белка [8].
Эндолизины частично ответственны за высвобождение новообразованных вирусных частиц за счет деградации PG, что дестабилизирует клеточную стенку и вызывает разрушение бактерии. Некоторые фаги также кодируют второй тип PGH, а именно вирион-ассоциированную пептидогликангидролазу (virion-associated peptidoglycan hydrolase — VAPGH), которая в отличие от эндолизинов разрушает PG во время начального проникновения бактериофага через клеточную стенку в бактерию. Геном фагов также содержит гены, кодирующие полисахаридные деполимеразы, которые деградируют полисахариды оболочки бактериальных клеток [53].
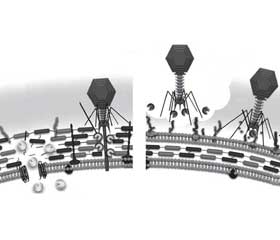
Эндолизины ответственны за деградацию PG клеточной стенки бактерий на поздних стадиях репликации литического фага, вызывая высвобождение новых образовавшихся вирусных частиц (рис. 1). Так как слой PG обеспечивает структурную целостность и жесткость бактериальной стенки, его разрушение может привести к гибели бактерии. Грамположительные бактерии, у которых отсутствует внешняя мембрана, высоко чувствительны к эндолизинам, в то время как для осуществления эндолизин-индуцированного лизиса грамотрицательных бактерий необходимо предшествующее влияние факторов, которые катализируют слияние внутренней и наружной мембран [42].
Сведения о генетически модифицированных энзибиотиках представлены в базе данных GMEnzy (http://biotechlab.fudan.edu.cn/database/gmenzy/).
Антистрептококковые бактериофаги и эндолизины
В настоящее время идентифицированы два бактериофага (Cp-1 и Dp-1), специализированных на киллинге бактерий Streptococcus pneumoniae [31]. Эффективность применения эндолизинов данных бактериофагов продемонстрирована при лечении различных инвазивных форм Streptococcus pneumoniae-инфекций, в том числе и при пневмониях (табл. 2).
/705-1.jpg)
Результаты исследования лечебного эффекта Cpl-1 показали, что внутривенное введение препарата не является оптимальным методом доставки антистрептококового эндолизина. Внутривенное введение в дозе 10 мг/кг с последующей непрерывной инфузией 5 мг/кг/ч Cpl-1 в течение 6 ч крысам со стрептококковой инвазивной инфекцией не приводило к благоприятному исходу заболевания. И только после болюсного введения 250 мг/кг с последующей непрерывной инфузией 250 мг/кг/ч –Cpl-1 в течение 6 часов наблюдалось снижение уровня бактериемии [14]. Ограниченный терапевтический эффект внутривенного введения эндолизина, вероятно, обусловлен коротким периодом полувыведения препарата. Согласно данным исследований эффективности лечения пневмококковых инфекций у животных, эндолизины устраняют бактериальную нагрузку при назначении сразу после заражения, до клинической манифестации болезни. Назначение в первые сутки после заражения одной внутрибрюшинной инъекции Cpl-1 мышам, страдающим тяжелой пневмококковой пневмонией, сопровождается 100% выживаемостью. По сравнению с эффективностью терапии амоксициллином (86 % выживаемости) Cpl-1 обеспечивал достоверно более высокий уровень выживаемости. Однако начало лечения со вторых суток после инфицирования сопровождается снижением вероятности выживания до 42 % [54]. Ингаляционное однократное введение эндолизина Cpl-1 в дозе 0,4 мг (в 25 мл физиологического раствора) при помощи MicroSprayer мышам, больным тяжелой стрептококковой пневмонией, способствовало предупреждению летального исхода заболевания в 80 % случаев. Через 10 суток после введения Cpl-1 у мышей отмечается значимое снижение бактериальной нагрузки в легочной ткани, в то время как у выживших мышей, не леченных Cpl-1, наблюдается увеличение числа КОЕ бактерий Streptococcus pneumoniae в легких. Однако применение только ингаляционного введения эндолизина недостаточно для полной элиминации бактерий Streptococcus pneumoniae [12].
Антистафилококковые бактериофаги и эндолизины
В настоящее время проводятся испытания (фаза I) двух препаратов, основанных на антистафилококковых фагах, P128 GangaGen и CF-301 Contrafect, которые предлагают использовать для деколонизации носоглотки от бактерий Staphylococcus aureus [50].
Результаты исследований муреин/пептидогликановых гидролаз, продуцируемых бактериофагами, позволяют считать, что данные энзимы могут быть использованы в качестве новых антибактериальных средств для борьбы с грамположительными патогенами. Ruth Keary и соавт. [26] установили, что бактерио–фагальная муреиновая пептидаза CHAPK вызывает быстрый лизис нескольких штаммов MRSA и предотвращает образование стафилококковой биопленки, не сопровождаясь проявлением побочных эффектов у экспериментальных животных. Различные штаммы обладают разной чувствительностью к эндолизинам специфических бактериофагов (табл. 3).
Эффективность применения эндолизинов данных стафилококковых бактериофагов продемонстрирована при лечении различных заболеваниий, индуцированных Staphylococcus aureus и MRSA (табл. 4).
/707-1.jpg)
Введение эндолизинов приводит к уменьшению уровня бактериемии Staphylococcus aureus и предотвращает летальный исход септического процесса у грызунов. Так, внутрибрюшинное введение эндолизина MV-L в дозе 0,05 мг (2 мг/кг) через 30 минут после системного заражения бактериями MRSA сопровождается выживанием 100% инфицированных мышей. Однако введение MV-L через 1 час после инфицирования сопровождается выживаемостью только 60 % инфицированных мышей [40]. Применение в терапии более мощного антистафилококкового эндолизина LysGH15 с исключительной антистафилолитической активностью, высокоспецифичной для штаммов MRSA, в той же дозе (2 мг/кг) способствует предотвращению летального исхода заболевания и при его введении через один час после инфицирования. Однако назначение LysGH15 не способствует полной санации крови. Терапия эндолизином LysGH15 приводит и к снижению уровня активности воспалительного процесса, в частности, к уменьшению концентрации IL-6, IFN-g [18]. Назначение LysGH15 сопровождается специфическим антительным ответом на данный эндолизин, но антитела не влияют на терапевтическую эффективность LysGH15 [56]. Mathias Schmelcher и соавт. [44] продемонстрировали, что применение эндолизинов LysK, phi11, 80α, 2638A или WMY при стафилококковой бактериемии предотвращает летальность у 100 % экспериментальных животных, что практически сравнимо с эффективностью сочетанной терапии лизостафином и ванкомицином. В то же время необходимо отметить, что терапия эндолизинами Twort и phiSH2 сопровождалась развитием летального исхода в 40 % случаев, а Р68 — не предотвращала летального исхода.
Антиклебсиеллезные бактериофаги и эндолизины
Основываясь на результатах исследований антибактериальной активности бактериофагов, Eun-Ah Park и соавт. [37] показали, что бактериофаги PKO111 и PKP126 могут быть хорошими кандидатами для разработки терапевтических средств с выраженной бактерицидной активностью по отношению к бактериям Klebsiella. Также миовирусы KP15 и KP27 субсемейства Tevenvirinae продуцируют холин, антихолин, спапин и эндолизин, который, собственно, таргетно разрушает PG бактерий Klebsiella. Установлено, что эндолизин KP27 характеризуется энзиматической специфичностью, стабильностью и отсутствием токсичности по отношению к эпителиальным клеткам человека [30].
Антисинегнойные бактериофаги и эндолизины
В настоящее время идентифицировано несколько бактериофагов (E79, JG024, PaP1, MPK1, MPK6, PAK-P1, M4, LUZ7 и PIK), которые показали эффективность при лечении экспериментальных инфекций, вызванных бактериями Pseudomonas aeruginosa, у животных [10, 20, 23, 27, 38, 47].
Краткая характеристика исследований, посвященных изучению эффективности терапии бактериофагами экспериментальных пневмоний, вызванных бактериями Pseudomonas aeruginosa, представлена в табл. 5.
Продемонстрировано, что интраназальное введение в респираторный тракт бактериофага PAK-P1 эффективно предотвращает поражение легких и предупреждает развитие неблагоприятного исхода летальной инфекции, вызванной бактериями Pseudomonas aeruginosa у мышей. Эффективность снижения бактериальной нагрузки в ткани легкого при применении бактериофага PAK-P1 у мышей с синегнойной пневмонией сочетается с уменьшением уровня провоспалительных цитокинов. Через 48 часов после введения PAK-P1 уровни концентрации IL-6 и TNF-α возвращаются к исходным значениям [10]. Laurent Debarbieux и соавт. [10] подчеркивают, что достижение 100% выживаемости у инфицированных животных отмечается только тогда, когда лечение бактериофагом PAK-P1 проводится не позже, чем через 2 часа после инфицирования бактериями Pseudomonas aeruginosa. Eric Morello и соавт. [33] показали, что интраназальное введение бактериофагов PAK-P или P3-CHA мышам с летальной пневмонией, вызванной бактериями Pseudomonas aeruginosa, сопровождается выживаемостью экспериментальных животных в 90–100 % случаев. Также установлено, что бактериофаг P3-CHA оказывает профилактическое влияние: назначение P3-CHA за 4 дня до инфицирования в 100 % случаев предупреждает развитие инфекционного процесса.
Возможность применения эндолизинов бактериофагов была продемонстрирована несколькими группами исследователей (табл. 6).
Catherine Paradis-Bleau и соавт. [36] продемонстрировали, что gp144 фага phiKZ представляет собой литическую трансгликозилазу, способную взаимодействовать с дезорганизованными бактериальными мембранами и приводить к гибели бактерии Pseudomonas aeruginosa. Также показано, что эндолизины Art-175, EL188, OBPgp279, LysPA26 обладают бактерицидной активностью против бактерии Pseudomonas aeruginosa [3, 4, 6, 19, 51]. Представляет интерес, что эндолизин LysPA26 может эффективно приводить к гибели не только бактерии Pseudomonas aeruginosa, но и бактерии Klebsiella pneumoniae и Escherichia coli. Однако эндолизин LysPA26 неэффективен против таких грамположительных бактерий, как Staphylococcus aureus [19].
Необходимо отметить, что эффективность бактериофагальной терапии пневмоний зависит от времени введения препарата относительно момента инфицирования и дозы введенного бактериофага. Практически все исследователи продемонстрировали, что максимальный антибактериальный эффект фаговых препаратов проявляется при их введении в первые два часа после инфицирования причинно-значимым патогеном.
Выводы
Таким образом, литические бактериофаги и эндолизины способны вызывать гибель антибиотикорезистентных бактерий, в связи с чем их использование в качестве бактерицидных лекарственных средств является потенциально эффективным терапевтическим методом противомикробной терапии. Высокий уровень эффективности бактерицидного действия эндолизинов позволяет предположить, что будущие лекарственные средства, созданные на молекулярной основе эндолизинов, могут стать ключевыми противомикробными препаратами при лечении инвазивных инфекций.
Конфликт интересов. Авторы заявляют об отсутствии какого-либо конфликта интересов при подготовке данной статьи.
Список литературы
1. Ackermann H.W. 5500 Phages examined in the electron microscope // Arch. Virol. 2007 Feb; 152(2): 227-43. doi: 10.1007/s00705-006-0849-1.
2. Alemayehu D. Bacteriophages φMR299-2 and φNH-4 can eliminate Pseudomonas aeruginosa in the murine lung and on cystic fibrosis lung airway cells / D. Alemayehu, P.G. Casey, O. McAuliffe et al. // MBio. 2012 Mar 6; 3(2): e00029-12. doi: 10.1128/mBio.00029-12.
3. Briers Y. Analysis of outer membrane permeability of Pseudomonas aeruginosa and bactericidal activity of endolysins KZ144 and EL188 under high hydrostatic pressure / Y. Briers, A. Cornelissen, A. Aertsen et al. // FEMS Microbiol Lett. 2008 Mar; 280(1): 113-9. doi: 10.1111/j.1574-6968.2007.01051.x.
4. Briers Y. Art-175 is a highly efficient antibacterial against multidrug-resistant strains and persisters of Pseudomonas aeruginosa / Y. Briers, M. Walmagh, B. Grymonprez et al. // Antimicrob. Agents Chemother. 2014 Jul; 58(7): 3774-84. doi: 10.1128/AAC.02668-14.
5. Briers Y. Engineered endolysin-based "Artilysins" to combat multidrug-resistant gram-negative pathogens / Y. Briers, M. Walmagh, V. Van Puyenbroeck et al. // MBio. 2014 Jul 1; 5(4): e01379-14. doi: 10.1128/mBio.01379-14.
6. Briers Y., Walmagh M., Lavigne R. Use of bacteriophage endolysin EL188 and outer membrane permeabilizers against Pseudomonas aeruginosa // J. Appl. Microbiol. 2011 Mar; 110(3): 778-85. doi: 10.1111/j.1365-2672.2010.04931.x.
7. Buttimer C. Bacteriophages and Bacterial Plant Diseases / C. Buttimer, O. McAuliffe, R.P. Ross et al. // Front Microbiol. 2017 Jan 20; 8: 34. doi: 10.3389/fmicb.2017.00034.
8. Cisek A.A. Phage Therapy in Bacterial Infections Treatment: One Hundred Years After the Discovery of Bacteriophages / A.A. Cisek, I. Dąbrowska, K.P. Gregorczyk, Z. Wyżewski // Curr Microbiol. 2017 Feb; 74(2): 277-283. doi: 10.1007/s00284-016-1166-x.
9. Daniel A. Synergism between a novel chimeric lysin and oxacillin protects against infection by methicillin-resistant Staphylococcus aureus / A. Daniel, C. Euler, M. Collin et al. // Antimicrob Agents Chemother. 2010 Apr; 54(4): 1603-12. doi: 10.1128/AAC.01625-09.
10. Debarbieux L. Bacteriophages can treat and prevent Pseudomonas aeruginosa lung infections / L. Debarbieux, D. Leduc, D. Maura et al. // J. Infect. Dis. 2010 Apr 1; 201(7): 1096-104. doi: 10.1086/651135.
11. Díez-Martínez R. A novel chimeric phage lysin with high in vitro and in vivo bactericidal activity against Streptococcus pneumoniae / R. Díez-Martínez, H.D. De Paz, E. García-Fernández et al. // J. Antimicrob. Chemother. 2015; 70(6): 1763-73. doi: 10.1093/jac/dkv038.
12. Doehn J.M. Delivery of the endolysin Cpl-1 by inhalation rescues mice with fatal pneumococcal pneumonia / J.M. Doehn, K. Fischer, K. Reppe et al. // J. Antimicrob. Chemother. 2013 Sep; 68(9): 2111-7. doi: 10.1093/jac/dkt131.
13. Doss J. A Review of Phage Therapy against Bacterial Pathogens of Aquatic and Terrestrial Organisms / J. Doss, K. Culbertson, D. Hahn et al. // Viruses. 2017 Mar 18; 9(3). pii: E50. doi: 10.3390/v9030050.
14. Entenza J.M. Therapeutic effects of bacteriophage Cpl-1 lysin against Streptococcus pneumoniae endocarditis in rats / J.M. Entenza, J.M. Loeffler, D. Grandgirard et al. // Antimicrob Agents Chemother. 2005 Nov; 49(11): 4789-92. doi: 10.1128/AAC.49.11.4789-4792.2005.
15. Fenton M. Recombinant bacteriophage lysins as antibacterials / M. Fenton, P. Ross, O. McAuliffe et al. // Bioeng Bugs. 2010 Jan-Feb; 1(1): 9-16. doi: 10.4161/bbug.1.1.9818.]
16. Gilmer D.B. Novel bacteriophage lysin with broad lytic activity protects against mixed infection by Streptococcus pyogenes and methicillin-resistant Staphylococcus aureus / D.B. Gilmer, J.E. Schmitz, C.W. Euler, V.A. Fischetti // Antimicrob Agents Chemother. 2013 Jun; 57(6): 2743-50. doi: 10.1128/AAC.02526-12.
17. Grandgirard D. Phage lytic enzyme Cpl-1 for antibacterial therapy in experimental pneumococcal meningitis / D. Grandgirard, J.M. Loeffler, V.A. Fischetti, S.L. Leib // J. Infect. Dis. 2008 Jun 1; 197(11): 1519-22. doi: 10.1086/587942.
18. Gu J. LysGH15 reduces the inflammation caused by lethal methicillin-resistant Staphylococcus aureus infection in mice / J. Gu, J. Zuo, L. Lei et al. // Bioeng Bugs. 2011 Mar-Apr; 2(2): 96-9. doi: 10.4161/bbug.2.2.14883.
19. Guo M. A Novel Antimicrobial Endolysin, LysPA26, against Pseudomonas aeruginosa / M. Guo, C. Feng, J. Ren et al. // Front Microbiol. 2017 Feb 27; 8: 293. doi: 10.3389/fmicb.2017.00293.
20. Harper D.R., Enright M.C. Bacteriophages for the treatment of Pseudomonas aeruginosa infections // J. Appl. Microbiol. 2011 Jul; 111(1): 1-7. doi: 10.1111/j.1365-2672.2011.05003.x.
21. Hobbs Z., Abedon S.T. Diversity of phage infection types and associated terminology: the problem with "Lytic or lysogenic" // FEMS Microbiol Lett. 2016 Apr; 363(7). pii: fnw047. doi: 10.1093/femsle/fnw047.
22. Hojckova K., Stano M., Klucar L. phiBIOTICS: catalogue of therapeutic enzybiotics, relevant research studies and practical applications // BMC Microbiol. 2013 Mar 6; 13: 53. doi: 10.1186/1471-2180-13-53.
23. Hraiech S. Bacteriophage-based therapy in cystic fibrosis-associated Pseudomonas aeruginosa infections: rationale and current status / S. Hraiech, F. Brégeon, J.M. Rolain et al. // Drug Des Devel Ther. 2015 Jul 16; 9: 3653-63. doi: 10.2147/DDDT.S53123.
24. Instytut Immunologii i Terapii Doświadczalnej PAN (2015) Najczęstsze pytania. www.iitd.pan.wroc.pl/pl/OTF/Pytania.html. Accessed 10 June 2015.
25. Jado I. Phage lytic enzymes as therapy for antibiotic-resistant Streptococcus pneumoniae infection in a murine sepsis model / I. Jado, R. López, E. García et al. // J. Antimicrob. Chemother. 2003 Dec; 52(6): 967-73. doi: 10.1093/jac/dkg485.
26. Keary R. Characterization of a Bacteriophage-Derived Murein Peptidase for Elimination of Antibiotic-Resistant Staphylococcus aureus / R. Keary, M. Sanz-Gaitero, M.J. van Raaij et al. // Curr Protein Pept Sci. 2016; 17(2): 183-90. doi: 10.2174/1389203716666151102105515.
27. Krylov V.N. Bacteriophages of Pseudomonas aeruginosa: long-term prospects for use in phage therapy// Adv. Virus Res. 2014; 88: 227-78. doi: 10.1016/B978-0-12-800098-4.00005-2.
28. Loeffler J.M., Djurkovic S., Fischetti V.A. Phage lytic enzyme Cpl-1 as a novel antimicrobial for pneumococcal bacteremia // Infect Immun. 2003 Nov; 71(11): 6199-204. doi: 10.1128/IAI.71.11.6199-6204.2003.
29. Loeffler J.M., Nelson D., Fischetti V.A. Rapid killing of Streptococcus pneumoniae with a bacteriophage cell wall hydrolase // Science. 2001 Dec 7; 294(5549): 2170-2. doi: 10.1126/science.1066869.
30. Maciejewska B. Klebsiella phages representing a novel clade of viruses with an unknown DNA modification and biotechnologically interesting enzymes / B. Maciejewska, B. Roszniowski, A. Espaillat et al. // Appl. Microbiol. Biotechnol. 2017 Jan; 101(2): 673-684. doi: 10.1007/s00253-016-7928-3.
31. Mariano R. The interactome of Streptococcus pneumoniae and its bacteriophages show highly specific patterns of interactions among bacteria and their phages / R. Mariano, S. Wuchty, M.G. Vizoso-Pinto et al. // Sci Rep. 2016 Apr 22; 6: 24597. doi: 10.1038/srep24597.
32. McCullers J.A. Novel strategy to prevent otitis media caused by colonizing Streptococcus pneumoniae / J.A. McCullers, A. Karlström, A.R. Iverson et al. // PLoS Pathog. 2007 Mar; 3(3): e28.doi: 10.1371/journal.ppat.0030028.
33. Morello E. Pulmonary bacteriophage therapy on Pseudomonas aeruginosa cystic fibrosis strains: first steps towards treatment and prevention / E. Morello, E. Saussereau, D. Maura et al. // PLoS One. 2011 Feb 15; 6(2): e16963. doi: 10.1371/journal.pone.0016963.
34. Nelson D., Loomis L., Fischetti V.A. Prevention and elimination of upper respiratory colonization of mice by group A streptococci by using a bacteriophage lytic enzyme // Proc. Natl. Acad. Sci U S A. 2001 Mar 27; 98(7): 4107-12. doi: 10.1073/pnas.061038398.
35. Oliveira H. Molecular aspects and comparative genomics of bacteriophage endolysins / H. Oliveira, L.D. Melo, S.B. Santos et al. // J. Virol. 2013 Apr; 87(8): 4558-70. doi: 10.1128/JVI.03277-12.
36. Paradis-Bleau C. Peptidoglycan lytic activity of the Pseudomonas aeruginosa phage phiKZ gp144 lytic transglycosylase / C. Paradis-Bleau, I. Cloutier, L. Lemieux et al. // FEMS Microbiol Lett. 2007 Jan; 266(2): 201-9. doi: 10.1111/j.1574-6968.2006.00523.x.
37. Park E.A. Characterization and genome analysis of novel bacteriophages infecting the opportunistic human pathogens Klebsiella oxytoca and K. pneumoniae / E.A. Park, Y.T. Kim, J.H. Cho et al. // Arch Virol. 2017 Apr; 162(4): 1129-1139. doi: 10.1007/s00705-016-3202-3.
38. Pires D.P. Phage Therapy: a Step Forward in the Treatment of Pseudomonas aeruginosa Infections / D.P. Pires, D. Vilas Boas, S. Sillankorva, J. Azeredo // J. Virol. 2015 Aug; 89(15): 7449-56. doi: 10.1128/JVI.00385-15.
39. Preclinical safety evaluation of intravenously administered SAL200 containing the recombinant phage endolysin SAL-1 as a pharmaceutical ingredient / S.Y. Jun, G.M. Jung, S.J. Yoon et al. // Antimicrob. Agents Chemother. 2014; 58(4): 2084-8. doi: 10.1128/AAC.02232-13.
40. Rashel M. Efficient elimination of multidrug-resistant Staphylococcus aureus by cloned lysin derived from bacteriophage phi MR11 / M. Rashel, J. Uchiyama, T. Ujihara et al. // J. Infect Dis. 2007 Oct 15; 196(8): 1237-47. doi: 10.1086/521305.
41. Reindel R., Fiore C.R. Phage Therapy: Considerations and Challenges for Development // Clin. Infect. Dis. 2017 Mar 1. doi: 10.1093/cid/cix188.
42. Roach D.R., Donovan D.M. Antimicrobial bacteriophage-derived proteins and therapeutic applications // Bacteriophage. 2015 Jun 23; 5(3): e1062590. doi: 10.1080/21597081.2015.1062590.
43. Schmelcher M. Chimeric phage lysins act synergistically with lysostaphin to kill mastitis-causing Staphylococcus aureus in murine mammary glands / M. Schmelcher, A.M. Powell, S.C. Becker et al. // Appl Environ Microbiol. 2012 Apr; 78(7): 2297-305. doi: 10.1128/AEM.07050-11.
44. Schmelcher M. Evolutionarily distinct bacteriophage endolysins featuring conserved peptidoglycan cleavage sites protect mice from MRSA infection / M. Schmelcher, Y. Shen, D.C. Nelson et al. // J. Antimicrob Chemother. 2015 May; 70(5): 1453-65. doi: 10.1093/jac/dku552.
45. Sharma S. Bacteriophages and its applications: an overview / S. Sharma, S. Chatterjee, S. Datta et al. // Folia Microbiol (Praha). 2017 Jan; 62(1): 17-55. doi: 10.1007/s12223-016-0471-x.
46. To K.H., Young R. Probing the structure of the S105 hole // J. Bacteriol. 2014 Nov; 196(21): 3683-9. doi: 10.1128/JB.01673-14.
47. Torres-Barceló C. A window of opportunity to control the bacterial pathogen Pseudomonas aeruginosa combining antibiotics and pha–ges / C. Torres-Barceló, F.I. Arias-Sánchez, M. Vasse et al. // PLoS One. 2014 Sep 26; 9(9): e106628. doi: 10.1371/journal.pone.0106628.
48. Trudil D. Phage lytic enzymes: a history // Virol Sin. 2015 Feb; 30(1): 26-32. doi: 10.1007/s12250-014-3549-0.
49. Vouillamoz J. Bactericidal synergism between daptomycin and the phage lysin Cpl-1 in a mouse model of pneumococcal bacteraemia / J. Vouillamoz, J.M. Entenza, M. Giddey et al. // Int. J. Antimicrob Agents. 2013 Nov; 42(5): 416-21. doi: 10.1016/j.ijantimicag.2013.06.020.
50. Vuong C. Investigational drugs to treat methicillin-resistant Staphylococcus aureus / C. Vuong, A.J. Yeh, G.Y. Cheung, M. Otto// Expert Opin. Investig. Drugs. 2016; 25(1): 73-93. doi: 10.1517/13543784.2016.1109077.
51. Walmagh M. Characterization of modular bacteriophage endolysins from Myoviridae phages OBP, 201φ2-1 and PVP-SE1 / M. Walmagh, Y. Briers, S.B. dos Santos et al. // PLoS One. 2012; 7(5): e36991. doi: 10.1371/journal.pone.0036991.
52. Weinbauer M.G. Ecology of prokaryotic viruses // FEMS Microbiol Rev. 2004 May; 28(2): 127-81. doi: 10.1016/j.femsre.2003.08.001.
53. Wittekind M., Schuch R. Cell wall hydrolases and antibiotics: exploiting synergy to create efficacious new antimicrobial treatments // Curr. Opin. Microbiol. 2016 Oct; 33: 18-24. doi: 10.1016/j.mib.2016.05.006.
54. Witzenrath M. Systemic use of the endolysin Cpl-1 rescues mice with fatal pneumococcal pneumonia / M. Witzenrath, B. Schmeck, J.M. Doehn et al. // Crit. Care Med. 2009 Feb; 37(2): 642-9. doi: 10.1097/CCM.0b013e31819586a6.
55. Wu H. GMEnzy: a genetically modified enzybiotic database / H. Wu, J. Huang, H. Lu et al. // PLoS One. 2014 Aug 1; 9(8): e103687. doi: 10.1371/journal.pone.0103687.
56. Zhang L. LysGH15 kills Staphylococcus aureus without being affected by the humoral immune response or inducing inflammation / L. Zhang, D. Li, X. Li et al. // Sci Rep. 2016 Jul 7; 6: 29344. doi: 10.1038/srep29344.

/703-1.jpg)
/704-1.jpg)
/705-1.jpg)
/706-1.jpg)
/707-1.jpg)
/708-1.jpg)